ТИРОНЕТ – все о щитовидной железе Для специалистов Журнал Тиронет Архив журнала 2001 год № 3
Резистентность к тиреоидным гормонам
О. Баккер, В.М. Версинга Амстердам (Нидерланды)
Division of Endocrinology, Academic Medical Centre, Meibergdreef 9, 1105 AZ Amsterdam,
The Netherlands Thyroid international 3 - 2001
Русский перевод к.м.н. Фадеева В.В. (примечания переводчика отмечены *)
ВВЕДЕНИЕ
В ряде случаев в клинической практике встречаются пациенты, у которых можно видеть симптомы напоминающие таковые при гипотиреозе или тиреотоксикозе в сочетании с противоречивыми результатами гормонального исследования: у них, несмотря на повышенный уровень Т4, может не выявляться подавления уровня ТТГ, а в ряде случаев он оказывается даже слегка повышен. Эту парадоксальную ситуацию может объяснять множество причин. Одними из них являются ТТГ-продуцирующая аденома гипофиза и резистентность к тиреоидным гормонам (РТГ). В этом обзоре мы обсудим последнюю патологию. В основе патогенеза РТГ лежит снижение чувствительности тканей к тиреоидным гормонам. Снижение чувствительности к тиреоидным гормонам гипофиза приводит к нарушению отрицательной обратной связи регуляции функции щитовидной железы (рис. 1), которое приводит к усилению продукции ТТТ несмотря на нормальный уровень Т4. Как будет показано дальше, нарушение чувствительности к тиреоидным гормонам связано с мутацией b-изоформы рецептора тиреоидных гормонов, что сопровождается снижением аффинности рецептора. Все это может приводить не только к появлению симптомов гипотиреоза, но и у некоторых пациентов, к развитию ряда симптомов тиреотоксикоза, в первую очередь тахикардии.
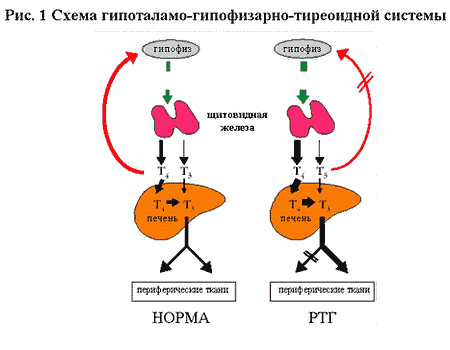
В норме гипофиз продуцирует ТТГ, который стимулирует продукцию Т4 щитовидной
железой. В печени Т4 метаболизируется до Т3 и оба эти гормона замыкают петлю отрицательной
обратной связи, подавляя продукцию ТТГ. Биологически наиболее активный Т3 высвобождается
в системный кровоток, достигает периферических тканей и активирует свои рецепторы.
При RTH вследствие снижения чувствительности гипофиза к тиреоидным гормонам нарушается
отрицательная обратная связь, в результате чего происходит неадекватное уровню
Т4 и Т3 повышение продукции ТТГ, который, в свою очередь, дополнительно стимулирует
щитовидную железу, обуславливая ги-перпродукцию Т4. Избыток тиреоидных гормонов
в организме на фоне RTH приводит к тому, что у пациента может выявляться сочетание
симптомов тиреотоксикоза и гипотиреоза, в связи с тем, что периферические ткани
могут обладать разной чувствительностью к тиреоидным гормонам - от полной резистентность
до нормальной чувствительности.
В норме окончательные эффекты тиреоидных гормонов на тканевом уровне обусловлены изменением экспрессии Т3-чувствительных генов (T3-responsive genes) и синтеза их продуктов. Например, увеличение уровня холестерина ЛПНП, которое обнаруживается при гипотиреозе, может быть связано со снижением экспрессии гена рецептора ЛПНП. Этот ген чувствителен к тиреоидным гормонам. Эффекты гормонов щитовидной железы опосредованы ядерными рецепторами тиреоидных гормонов (TR), которые существуют как минимум в 4 изоформах (рис. 2). TR являются членами так называемого семейства ядерных рецепторов, к которому, кроме того, относятся рецепторы стероидных гормонов, витамина Д и ретиноевой кислоты. Эти рецепторы изменяют эксперссию генов за счет связывания со специфическими элементами ДНК, которые обозначаются как тиреоид-чувствительные элементы (TRE - thyroid response elements) и локализованы в промоторном регионе Т3-чувствительных генов. TR могут вступать в эту связь в виде гомодимера (два идентичных мономера) или гетеродимера (два разных мономера). Выделяют три активные формы TR, а именно TRa1, TRb1 и TRb2, которые связывают Т3 и одна неактивная - TRa2, которая не связывает Т3. Все они являются производными двух различных генов, которые локализуются на хромосомах 17 и 3 соответственно и по-разному экспрессируются в различных тканях (рис. 2). Кроме того, TR могут связываться с различными TRE, в ряде случаев в специфических комбинациях, что свидетельствует о наличии большого числа специфических регуляторных эффектов на различные функции. Это объясняет тот факт, что TR (после связывания с Т3) может как активировать гены (например, ген рецептора ЛПНП), так и ингибировать их. Примером подавления активности генов может быть отрицательная обратная связь продукции ТТГ и тиреоидных гормонов, когда связанные с лигандом тиреоидные гормоны присоединяются к специфическому TRE генов a- и b-субъединицы ТТГ, что приводит к подавлению активности этих генов.
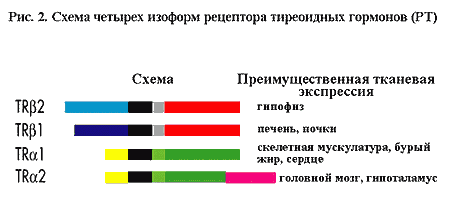
Схематически представлены различные изоформы рецепторов тиреоидных гормонов
(РТ) с указанием тканей, где они преимущественно экспрессирова-ны. Черным представлен
ДНК-связывающий домен, красным и темно-зеленым - Т3-связывающие домены. TR?1 и
TR?2 отличаются N-терминальными доменами, тогда как TR?1 и TR?2 - С-терминальными.
В обоих случаях мРНК транскрибируется с одного и того же гена, который в результате
альтернативного сплайсинга приобретает a или b изоформы.
Рецепторы тиреоидных гормонов являются уникальными представителями своего семейства в том плане, что они могут изменять экспрессию генов как при наличии лиганда, так и без него. Это связано с тем, что TR может связываться с TRE при отсутствии гормона. В последнем случае TRE инициирует так называемый белок корепрессор, который подавляет активность гена. Среди корепрессоров в настоящее время известны такие соединения как N-CoR и SMRT [Zhang и Lazar, 2000]. В присутствии тиреоидных гормонов рецепторный гомодимер освобождает корепрессор, после чего распадается на мономеры. Связанный с лигандом рецепторный мономер образует гетеродимер с ретиноевым Х рецептором (RXR) и, несмотря на то, что присоединяется к тому же TRE, благодаря другим конформационным изменениям, он приобретает способность инициировать коактиватор, который в свою очередь, значительно усиливает транскрипцию ряда генов (рис. 3). К настоящему времени описано несколько коактиваторов TR, в частности SRC-1 и TIF2/GRIP1 [Zhang и Lazar, 2000].
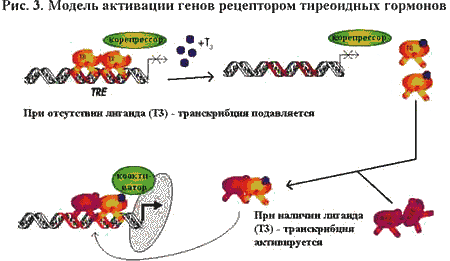
Когда TR не связан с лигандом, он соединяется с TRE в виде димера (два идентичных
мономера TR) и подавляет экспрессию гена за счет взаимодей-ствия с корепрессором.
Если гормон связан с TR, корепрессор высвобожда-ется, а гомодимер TR распадется
на мономеры. Мономер TR вступает в гете-родимерзацию с ретиноевым Х рецептором
(RXR), после чего гетеродимер вновь связывается с TRE, инициирует коактиватор,
который стимулирует транскрипцию гена.
Недавно были охарактеризованы три пространственные структуры рецепторов тиреоидных гормонов [Wagner, 1995]. После этого стало очевидным, что тиреоидные гормоны в результате взаимодействия с рецепторами оказываются глубоко «запакованы» в глубине их молекулы и что последние несколько молекул аминокислот рецепторного белка (двенадцатая спираль) действуют как своего рода крышка, которая закрывает «ящик» куда попал гормон. Результаты этих исследований кроме того показали, что «закрытие крышки» необходимо для конформационного формирования связывающего участка, к которому присоединяется коактиватор.




